01/09/08
Feeding in Aquatic Organisms
We have considered many aspects of living in water, from the physical and chemical basics to the topics of osmoregulation, respiration, locomotion and sensory perception. We have discussed how plants obtain the dissolved gasses and inorganic nutrients that they need for photosynthesis; now it is time to consider how animals obtain the chemicals they need for life.
There are two basic sources of food for aquatic organisms. Photosynthesis uses energy from the sun to incorporate CO2 into carbohydrates, which animals can break down for energy; chemosynthesis uses the energy inherent in reduced metal compounds for the same purpose. Photosynthesis in water is a surface phenomenon; organisms in the deep cannot use photosynthesis directly. Chemosynthesis usually requires an anoxic environment, and in the modern world is limited to certain small, scattered habitats.
Plants can be divided into the algae, which are small, easy to ingest, relatively diffuse in the environment, and relatively hard to digest. Higher plants are large, hard to ingest, and hard to digest, and in any event, are relatively rare in aquatic habitats. Plants in general are a food of intermediate quality. Animals are mobile, they may fight back, and therefore are overall usually hard to ingest, but once ingested they provide high-quality food that is easy to digest. Detritus, which consists of decaying plants, animals, and animal waste, comes in a variety of sizes, is usually easy to ingest, is usually easy to digest, but is also of the lowest quality.
To summarize, animal material is of the highest quality, is the easiest to digest, but is the toughest to ingest. Plants are of medium quality, are easy to get, but may be tough to ingest, and are almost always tough to digest. Detritus is the easiest to ingest, is of medium digestibility, but has the lowest quality. Of course, these are generalizations; for instance, a recently killed animal is technically detritus but is of far above average quality. The food source for an organism can be very important in its life cycle. For instance, mosquito larvae, which feed on low-quality detritus, often reach the adult stage without enough protein to produce eggs, thus forcing the females to seek out blood meals. Organisms that feed on decaying wood (mostly cellulose) often depend on symbionts or fungi to obtain protein. Plant feeders must overcome not only thick, indigestible cell walls, but also a host of chemical toxins stored by the plants to dissuade herbivores.
There are several ways to divide the continuum that animal feeding modes fall into up into discrete categories, and, after 10 years of research into how aquatic insects feed, I can tell you that about the only generalization that you can make about animal feeding habits is that most animals are opportunistic - they will eat whatever is available. There are specialists, and there are organisms that specialize some of the time, but a lot of animals simply resist being classified as to feeding type, particularly if you observe them over their entire life cycle. This tendency seems to be especially true for many aquatic organisms, which tend to be generalists. Indeed, it might be argued that a high degree of specialization in feeding is largely a phenomenon confined to terrestrial organisms and parasites.
With that in mind, we will nevertheless attempt to categorize feeding in aquatic organisms. The next hurdle comes in trying to decide how to make the distinctions between feeding groups, that is, what criteria to use. There are several different criteria that can be used, and each reveals a different type of information. The choice of which system to use is thus largely a function of the type of information you wish to gather.
Dietary Feeding Groups
Often ecologists are interested in tracing the flow of energy through an ecosystem. From this standpoint, a system that keeps track of diet is valuable, particularly if that diet is related to what are known as trophic levels - where in the food chain the energy present in the prey came from. Such a system typically divides the world up as follows:
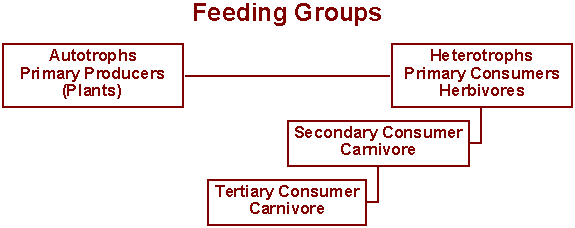
This system could be continued on through 4o and 5o consumers and so on until the end of the food chain is reached. Another, similar system is also included above and includes the old favorites herbivores and carnivores. The advantage of this system is that you can make predictions of population size and energy flow if all the organisms stay in their groupings. For instance, a general rule of thumb for food chains is that only about 10% of the energy at one level is available to be transferred to the next. So, if all your organisms have about the same biomass, you would expect to have about 10x as many plants as primary consumers, 10x as many primary consumers as secondary consumers, and so on.
The big problem with this type of system is that organisms do not always fall into such neat categories. Some animals are omnivorous, eating both plants and animals; some animals are detritivores, feeding on decaying plant and animal remains or wastes, often without regard to the origin of the material, and it is unrealistic to expect a hawk to think "gee, I've been classified as a tertiary consumer, so I better eat only secondary consumers, and that means asking this snake to be sure she's only been eating primary consumers, so that when I eat her I don't inadvertently become a 4o consumer ...". In other words, animals ignore the boundaries and the ecologist must determine what percent of the animal's nutrition comes from each of the other trophic levels. This makes things a great deal more complicated.
There are other diet-based classifications as well; some of which work pretty well for specialists that concentrate on a particular type of food. For instance, we can speak of piscivores (fish eaters), insectivorous plants, frugivores (fruit eaters), or even coprovores (dung eaters).
Functional Feeding Groups
An alternative to a diet-based classification is one based on how the food is ingested. These systems are sometimes known as functional feeding groups or FFG's. These FFG's arise almost ad hoc as workers describe feeding modes for organisms they are studying, without regards for what workers dealing with other groups of animals are doing. FFG's are based on physical characteristics of the food. I would suggest the following four continua as the characteristics by which food can be classified:
- Size - micro to macro (relative to organism feeding)
- Degree of attachment - suspended to deposited to attached
- Origin - protist to plant to animal
- Condition - living to dying to decomposing
Note that the diet-based classification scheme is based on the last two continua, origin and condition. FFG's are usually based more on the first two, size and degree of attachment.
One fairly good system for aquatic insects was developed originally by Merritt and Cummins (1978). It was greatly improved by a paper that came out a decade later (McShaffrey and McCafferty, 1988). Before we get into the details, though, a little background is needed.
There are several sources of food for aquatic insects. These sources include algae, both attached (periphyton) or as plankton, other insects, fish, vascular plants, and detritus. This food can be further divided as to size, either microscopic or macroscopic, and degree of attachment to the substrate, from suspended in the water (seston) to loosely deposited on the substrate to actually attached. Traditionally, the feeding categories in Merritt and Cummins were assigned after looking at the gut contents of the organisms in question. This is a risky business because gut contents are hard to identify and, since the organisms are so opportunistic, gut contents may vary from day to day (or season).
A much better way to determine what an animal is eating is to actually watch what the animal eats, a common-sense approach rarely employed in the past. To be fair, it is extremely difficult to observe feeding under natural conditions; these are, after all, small organisms, living underwater, often in flowing, turbid waters, often in the dark under rocks. Furthermore, accurate determination of the function of different feeding structures is impossible without detailed movement analyses and magnifications of the feeding structures above that possible with a light microscope Scanning electron microscopy solved the second problem, and inexpensive consumer video equipment, which began to appear in the 1980's, solved the first.
By combining these tools, I was able to observe feeding in detail, and correlate feeding structures and feeding habits. With this data now available, it is possible for me to make fairly accurate predictions as to what an organism eats and where it lives simply by examining its structure (people at scientific meetings often bring me pictures and ask me "what does this thing eat?"). The feeding scheme I devised along with Dr. W.P. McCafferty modifies the Merritt and Cummins scheme by including this new data and providing functional (as opposed to dietary) definitions for some of the groups defined by Merritt and Cummins. The McShaffrey-McCafferty scheme works for microvores (those which feed on microscopic particles) among the aquatic insects (and also among many other benthic aquatic organisms) and is stated as follows (from McShaffrey and McCafferty, 1988):
"The ability of an organism to use more than one mode of feeding, whether it be R. pellucida scraping and brushing, or S. interpunctatum using the same mouthparts to brush, collect-gather, or filter; or different developmental stages of a single species using different feeding methods (Cummins 1973, Cummins and Merritt 1984), complicates the problem of pigeonholing species into FFG's. This complexity is aggravated by attempting to assign FFG's based on gut contents that are often unidentifiable. Our study of R. pellucida and other aquatic insects suggests that a more mechanical approach to delineating FFG's for benthic insects that feed on relatively small materials (microvores, sensu McCafferty 1981) may be appropriate. In this scheme, potential food material, regardless of origin, is viewed in terms of a continuum ranging from material suspended in the water, to material settled on the substrate, to material bound or growing attached to the substrate. Various feeding strategies are adapted to deal with various sections of this continuum.
In our scheme, benthic microvores can be divided into two basic groups, Filterers and Collectors. Filterers derive food material from the water, and the filter may consist of either parts of the body or manufactured devices such as silk nets. Passive filterers rely on seston already moving to their filtering apparatus, whereas active filterers generally resuspend deposits to filter. Thus, depending on the immediate source of the food material, we classify benthic filterers as either seston filterers, or deposit filterers. Collectors remove deposits or attached material from a substrate by direct contact of the feeding structures with the food material. Collectors can be divided into three groups: brushers, gatherers, and scrapers. Brushers use setae to obtain loose or lightly attached material and are often morphologically, functionally and behaviorally close to deposit filterers. Gatherers feed on similar materials but primarily use structures other than setae for food gathering. Scrapers have adaptations that allow them to feed on tightly accreted material.
To summarize, benthic filterers and collectors exhibit a range of feeding strategies for feeding on a spectrum of small particles ranging from suspended materials (seston filterers) to deposits of various integrity (deposit filterers, brushers, gatherers) to firmly attached materials (scrapers). As is the case for R. pellucida and S. interpunctatum, species may not be limited to one strategy. This scheme is attractive because habitats can be characterized hydraulically, and the FFG composition of the microvore community can be estimated based on the physical state in which the hydraulic forces will place the food material. In this context, concepts of community development can be based on the relative amounts of microhabitat available to provide food material in different positions in the environment and with different propensities for attachment. One drawback of such a classification system is that it provides little information about the origin of the material (i.e. primary production, detritus, etc.); however, under natural conditions many benthic macroinvertebrate microvores feed on a variety of materials and only a relative few species are dependent on a single trophic level for their food resources. As indicated above, the system excludes macrovores such as shredders, miners, engulfers, and many predators.
It remains premature to erect a new FFG classification system; however, such a system may become necessary as the behavior of more species of aquatic insects is studied in detail. For instance, Dahl et al. (1988) recently presented a classification scheme of FFG's for the Culicidae that also further divided the FFG's of Cummins and Klug (1979) into more discreet groupings based on the mechanism of food acquisition. It is notable that Dahl et al. (1988) chose to use the term brushers to describe a feeding system found in certain Culicidae. Those authors were evidently unaware of the brusher concept of McShaffrey and McCafferty (1986); from their discussion it is not clear whether their mosquito "brushers" feed by direct contact of the mouthparts with deposits (as is the case with S. interpunctatum), or if the mouthparts create a current which carries the material to the mouthparts (deposit filtering, herein), or both. "
With this feeding classification, which uses only the first two of our food classification continua (size and degree of attachment), there is little information on where the material originates, which is O.K., because the food in actuality originates in many places. For instance, a collector will be feeding on detritus, settled plankton, and animals simultaneously, defying classification in any one diet-based category. Because the FFG's are tied to the morphology of the animal, it is less likely (but still probable) that the animal will "break type", that is, a filter feeder spends most of its time filtering.
The McShaffrey-McCafferty scheme is probably too detailed for our use. As far as discussions of feeding go, we will use whatever scheme is appropriate. We will often talk in terms of dietary classifications, but for those times when a functional classification is more important, perhaps the scheme below will work:
- Microvores
- Suspension feeder (suspended material)
- Deposit feeder (unspecialized, loose, settled material)
- Grazer (attached material)
Macrovores
- Shredders
- Engulfers
- Piercers
Let us explore each of these groups in a little more detail (see also Fig. 1). Suspension feeders are the filterers of the McShaffrey-McCafferty scheme. They filter small particles from the water using a variety of techniques. Before we look at some examples, we must correct one bias that we have about filtering. Our concept, rooted in our high Re world is more of sieving than filtering. We see a filter as some sort of sieve, and the things retained by the sieve are those too big to fit through the holes. At low Re, that is only one possible mechanism. Smaller particles that you would expect to go through the filter may become entrapped due to gravitational forces (between the particle and the filter), electrostatic charges, or through what is known as inertial impaction, among others (Rubenstein and Koehl, 1977). What this means is that at low Re you can't judge what size particles will be filtered out simply by examining the mesh size - the size of the holes.
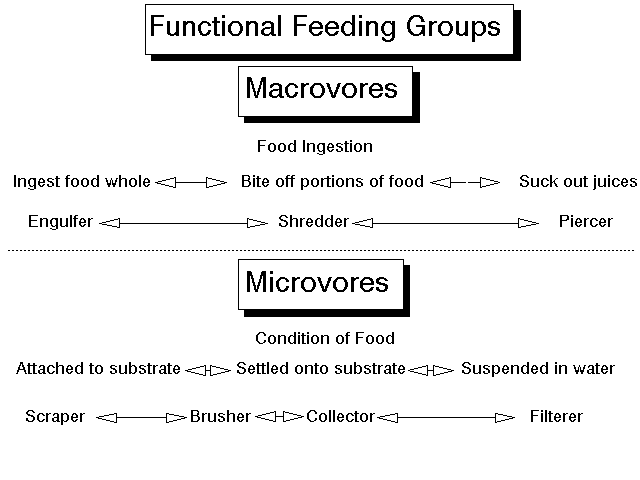
Figure 1. Functional Feeding Groups, shown here as functions of food size (macrovores) or degree of attachment of food to the substrate (microvores). Among the macrovores, the continuum encompasses the size of the prey as it is ingested, ranging from simply swallowing the prey whole (engulfer), to biting off pieces of the prey (shredder), to only ingesting the juices of the prey (piercer). Among the microvores, the continuum is drawn over degree of attachment of the food to the substrate, and ranges from tightly attached, to deposited, to suspended. In this scheme, scrapers remove tightly bound material; brushers remove more loosely bound material and cross over to material that has simply settled out; collectors remove deposited material; and filterers remove material from the water column.
As we have seen, filter feeders can be active, creating their own currents, or passive, letting water currents do the work. Let's examine some of the filtering structures used by animals. Sponges utilize the flagella of the collar cells to create a current; food settles out at the collar cell. Tunicates use water currents often generated by cilia to bring food to the pharynx, which is highly perforated to allow the water to pass, while the food is retained on a mucus coating. A similar scheme is used by bivalve mollusks such as clams. Lophophorates use mucus-covered tentacles in conjunction with ciliary currents to trap food; tunicates use the perforated pharynx as a filtering basket and in some species use the exhalant filtering current for propulsion. Some species use nets of mucus. Tentacles are also used by some filter-feeding echinoderms such as crinoids. Arthropods use either setae (hairs) for filtration, or they weave silk nets to filter material from the water. Many arthropods place the filtering structures on legs or other appendages where they can actively be moved through the water; such is the case for most of the planktonic crustaceans. Often the difference between a filter and a paddle is very slight, minute changes in the spacing of the setae or the speed of travel may make a huge difference (Rubenstein and Koehl (1977). Some filter feeders such as crinoids are little removed from deposit feeding cousins; their short stalk simply gets them above the substrate and gives them "first shot" at settling material. Remember that filter feeders include the largest organism that ever lived - the blue whale, which feeds by swallowing a huge mouthful of water and then spitting it out through highly modified teeth known as baleen. You can simulate the effect by taking a spoonful of cereal or soup and spitting it out through your teeth. Do this in front of young children and tell their parents you are teaching them about nature. The parents will love you.
Deposit feeders would be the gatherers and some of the brushers of the McShaffrey and McCafferty scheme. Some of them even cross the line a bit into filterers because they move to a deposit and stir it up and then filter out the food. Deposit feeders are generally feeding on detritus, dead and decaying animal or plant material that has fallen to the bottom. Coprophagy (eating feces) is also common among deposit feeders; some food items may end up passing through the guts of several animals. Since the material is loose and unattached, little specialization is needed in feeding structures. Any way to shove the food into the mouth will usually work; this even includes burrowing through the deposit with your mouth open. Many aquatic crustaceans use structures that resemble the filtering structures found on true filterers, and other organisms use mucus strands or nets in conjunction with cilia. At the other extreme, for material that is somewhat more cohesive, variations on the scraping strategies below will be employed. Brushers, for instance, are really a cross between a filter-feeder and a scraper.
Removing tightly bound material such as some diatoms or attached algae is a lot of work. Grazers can exploit this relatively (compared to detritus) high quality food where most deposit feeders cannot. Perhaps the most successful scraping structure in the animal kingdom is the radula of mollusks such as snails. Basically a series of small teeth that can be dragged over the surface to dislodge food, the radula is what separates the mollusks from the flatworms and is in large part responsible for the enormous success of the mollusks. Aquatic insects may duplicate the mechanical properties of the radula with specialized chitinous setae on their mouthparts, or with their claws. A researcher at Bowling Green and I are looking into the uncanny resemblance of snail and insect scraping structures. Other grazers might include fish such as the parrotfish which bites off pieces of coral and digests the polyps and associated organisms and expels the sand (much of the coral sand of the world starts out as parrotfish feces; think about that the next time you spread a towel). You should also be aware of the tendency of terrestrial ecologists to use the word grazer to refer to animals that feed by cropping grass and other short plants.
Organisms that feed on large pieces of food, macrovores, can be divided into three main groups depending on how they ingest the food. If they bite it off they are shredders; if they swallow it whole, they are engulfers; if they suck it out, they are piercers. Most vertebrates except lampreys and vampires are either engulfers or shredders. Engulfers and shredders will usually have teeth and jaws (or their equivalent) to facilitate feeding. Teeth may be sharp to hold prey before it is swallowed whole, or may be modified to cut or crush the prey. Herbivores in particular often have crushing mouthparts to break up tough cell walls and extract the contents; carnivores often have tearing or holding teeth. The beak of turtles, birds, cephalopods, and some fish is another tearing structure. Carnivore engulfers and shredders may employ specialized strategies and structures to capture prey (gory details later).
Macrovore piercers use a different strategy. They pierce the surface of their food and suck out its juices, often with the help of digestive enzymes. This is a common feeding method terrestrially, as anyone bothered by mosquitoes can attest. In aquatic systems it is also practiced by insects such as giant water bugs, by lampreys, and even by some mollusks such as clam larvae which are ectoparasitic on fish. Some aquatic insects even suck the juices out of plants in this way.
Another type of feeding which technically is a type of macrovore feeding is parasitism. In aquatic systems this will include miners; organisms (usually insects) which chew their way through the insides of plants sort of internal shredders). Animal parasites usually chew their way through the host tissue or simply absorb nutrients.
Evolution of Feeding Strategies
At this point, let's stop and consider how various feeding strategies may have evolved. The first multicellular animals (Fig. 2) probably ate each other, bacteria (including cyanobacteria), and protists. This diet seems limited until you consider that at the time of the first multicellular animals, that was all there was to eat. The descendants of these organisms that are still alive today are filter feeders (Porifera), carnivores (Cnidaria, Ctenophora), or deposit feeders (Placozoa, Mesozoa, and Platyhelminthes). The real story in terms of evolution of feeding strategies begins with the flatworms.
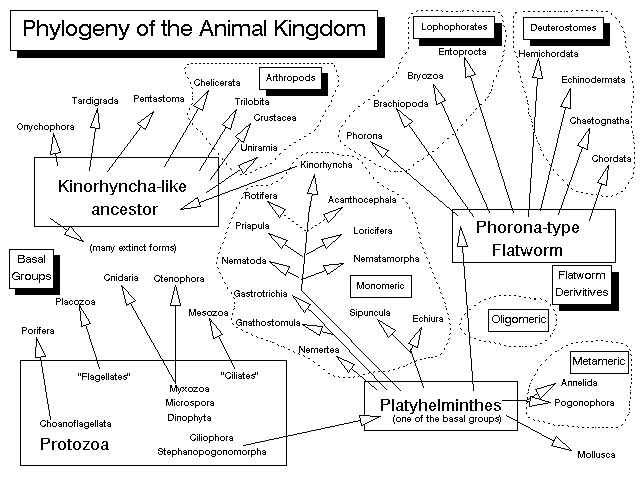
Figure 2. Phylogeny of the animal kingdom. See text for details on feeding habits as related to this phylogeny.
The basic flatworm is a deposit feeder that simply shovels stuff into its pharyngeal tube. Many of the phyla derived from the flatworms do likewise, with some important exceptions. The Mollusca have diversified so greatly partly in response to the many successful feeding strategies they have adopted, such as grazing with the use of the radula, predation, and filter feeding. Nematodes and some other groups have become adapted (in some cases) to live as parasites, although here their feeding movements are not really any different than their free-living forms, which are basically deposit feeders.
One type of flatworm, very close to today's Phorona, gave rise to the lophophorate and deuterostome phyla. These groups are mostly deposit or filter feeders, with the major exceptions being some predatory echinoderms and the myriad feeding groups found in the Chordata. Even within the chordates, however, early forms were primarily predators or deposit feeders; many of the feeding forms we see today evolved later, after the evolution of flowering plants. Organisms which evolved from the kinorhynch line also moved into many different feeding groups, primarily after the evolution of paired limbs which could be used for locomotion, prey capture, and as mouthparts.
In all these cases, the forces that seems to drive the development of different feeding groups from an unspecialized deposit feeder seem to be very similar. All are based in the fact that deposit feeders are ingesting a relatively low-quality food, and thus a switch to anything else is advantageous.
Animal tissue is the most nutritious food, and many organisms have adapted to live as carnivores. Among predators, there apparently are advantages to larger size. Larger organisms have a wider range of prey to select from, are less likely to be eaten themselves, and can bear more young. There is a thus a trend towards larger size among carnivores, and a constant development of accessory structures to find and capture prey (eyes, sharp teeth, claws, spines, etc.).
Plant tissue is also relatively nutritious, but hard to ingest and digest. These factors drive the selection of specialized feeding structures such as radulae or grinding teeth to help ingest the food, and a large gut with plenty of room for endosymbionts to help break down cellulose (animals in general don't seem to have evolved efficient cellulases yet). In aquatic systems, of course, most of the plant material ingested is either phytoplankton or algae scraped from rocks.
Among detritivores, the trend seems to be an effort to get the freshest detritus (and a mix of small living organisms if possible). In aquatic systems this means intercepting the material before it gets to the bottom. This factor encourages the development of stalked forms such as crinoids, lophophorates and brachiopods; large filtering structures such as poriferan colonies; and mobile filtering such as in whales, some tunicates, and others.
The development of terrestrial vascular vegetation also encouraged the development of successful strategies for ingesting such plants, and where vascular plants have reentered the water, terrestrial herbivores have soon followed. Examples of these would include a host of aquatic insects and even such large mammals as the manatee.
The Gut as it Relates to Feeding
The gut of most organisms can be divided into three functional regions - the foregut, the midgut, and the hindgut. Some organisms, however, do not have a through gut and thus will not have these regions; flatworms, Cnidaria, sponges and many parasites for instance do not have through guts of this sort.
In organisms with through guts, the divisions are both spatial and functional. The foregut usually consists of a mouth adapted for ingesting food. Often the mouth plays a very important role; the flexible cheeks of many fish and turtles help suck in prey. The mouth cavity may be the site of initial digestion if salivary glands are present. Chewing and tearing structures in the mouth may also begin physical digestion of the food. Posterior to the mouth is the pharynx, which in some organisms simply begins the process of swallowing; in others the pharynx forms a pump for sucking fluids into the mouth. Some fish such as the carp have pharyngeal teeth that also process the food as it is swallowed. Behind the pharynx is the esophagus used to connect the mouth with the crop, a food-storage organ. Organisms that feed only infrequently, such as several deep-sea predatory fishes, may have crops that can enlarge to a size greater than that of the entire body before eating.
The midgut begins with the stomach (often combined with the crop). The stomach's role is to initiate chemical digestion through the addition of secretions containing digestive enzymes from various digestive glands. The muscular stomach also continues physical digestion by churning the food along with the digestive juices. Some stomachs have teeth or grinding structures to further break up the food; stomachs of this sort are often called gizzards. Many organisms have various blind sacs or diverticula leading off the main digestive tract just posterior to the stomach; these structures provide more room for digestion and more surface area for absorption. Such structures are more common in herbivores. The intestine follows the stomach and is the site of nutrient absorption. It often is highly coiled, or has many internal folds, both ploys to increase surface area. As a general rule, the intestine of an herbivore will be longer and more coiled than that of a carnivore, since it takes more time (space) to remove nutrients from plants than animals, and because more plant material must be processed to obtain the same amount of nutrition.
The hindgut is the site of feces storage and water reclamation. The later process is, of course, more important to terrestrial organisms than aquatic ones, but remember that even in some aquatic organisms such as aquatic insects, chloride cells in the rectum are an important factor in osmoregulation.
The presence of cellulose and certain other molecules in food provides a definite challenge to digestive systems in animals. As mentioned before, few animals have developed any cellulase at all, and what is present is not very effective. Other molecules may be encountered too rarely to exert enough selective pressure for the development of specialized digestive enzymes for those molecules. The answers in both cases are symbionts, bacteria and protists that break down complex molecules in the gut and often make up bulk of feces. These symbionts often live in the diverticula of the digestive system, and, of course are more important to herbivores. They are widespread in aquatic organisms. Problems with symbionts include both initial inoculation of the gut and maintenance of a significant population in the forward reaches of the gut (since the movement in the gut is all "downstream"). The former (and often the latter) is often taken care of by coprophagy (eating dung); even in animals feeding on plants or attached algae some feces may be ingested. Mammal young probably pick up their inoculations from their mother or nestmates. If food tends to pass through the gut too fast for digestion or the maintenance of symbiont populations then autocoprophagy may result, where the organism eats its own feces.
Predation - Some More Details
The phenomenon of predation places some common selective pressure on organisms that feed in this way. Although there are many different ways to capture and/or feed on other organisms, the basic problems remain similar, and it is no surprise that similar solutions have evolved among carnivorous animals regardless of taxa. In order to feed on other organisms, they must be found, caught, killed, and ingested.
Finding prey often involves sophisticated sensory structures. All senses are used by predators in detecting potential prey. We are perhaps most familiar with sight, which, as it turns out, is perhaps the least used in water, since water is often dark and/or turbid. Still, there are a number of predators that use sight to some extent. These are primarily organisms that feed in shallow, clear habitats such as coral reefs, although our best local representatives might be centrarchids such as bass and bluegills. On land, predators are often told from herbivores by the presence of forward-directed eyes with good overlap of the visual field for stereoscopic vision, a necessity in gauging distance to the prey. Herbivores usually have lateral eyes with wide peripheral vision. In aquatic systems, however, this system breaks down, with only the top predators, such as alligators, having much in the way of stereoscopic vision. Other predators, such as bass, are simply too vulnerable to predation themselves, particularly when young.
Sound and related disturbances in the water are frequently used by predators to find prey. We are of course familiar with hearing as a tool in locating prey, but of course fish also have the lateral line to help them sense disturbance in the surrounding water. Marine mammals often use sound waves that they themselves generate to locate prey by bouncing the sound waves off the prey. This sonar is a highly advanced sense, and porpoises at least are able to discriminate between objects very close in size with their sonar. Their sonar sense, in fact, may have better resolution than vision under all but the clearest conditions. The sound waves that are generated have a short wavelength and are above the normal hearing range for humans. Many marine mammals have a "lens" of fat or oil in their head that serves to focus the sound waves, making this sense highly directional as well. The use of sound to locate prey is not limited to vertebrates, either. Recent evidence (Peckarsky and Wilcox, 1989) indicates that stoneflies can "hear" the swimming movements of their mayfly prey.
Perhaps the best distance sense in water is olfaction. Many predators can home in on their prey by scent alone, and, of course, many prey learn to avoid areas where the scent of potential predators is strong. Olfactory senses are strong among most aquatic organisms with the possible exception of aquatic insects, which perhaps have not developed these senses to their fullest potential yet. Apparently, the great feats of homing attributed to salmon, eels, and sea turtles in returning to the river (or beach) of their birth to mate and reproduce is due in part to an ability to chemically distinguish between different sites. Sharks have been known to follow blood trails of their prey for miles, and crayfish in tests have responded to vanishingly small dilutions of extracts from their food sources. Of course, as a distance sense, olfaction in water is at the mercy of the currents; for instance, in a stream a predator cannot detect the scent of a prey organism just slightly downstream, while it may be able to detect other prey far upstream. Similarly, turbulence in the water may either confine the scent to a narrow, concentrated stream or spread it into a wide, diffuse band. The former makes it easy to home in on the prey once the stream is detected, but the chance of detecting the odor is slight; in the latter, it is easy to detect the scent, but hard to home in on it to find the prey.
Other senses are commonly used in water as well. Perhaps the most important of these is the electric sense, the ability of specialized cells to detect the weak electrical energy given off by all living things. Because water conducts electricity, this sense is particularly well developed among aquatic organisms, especially those living in murky water. The ability to exploit electrical senses is found in both vertebrates and invertebrates, but is best studied in the vertebrates, where it is found in all major groups. The duck-billed platypus, for instance, uses electrical sensors in its bill to locate food in the mud. Sharks use the electrical sense to make the final biting lunge for their prey; at close range the smell of the prey is overpowering, and the eyes may close (or the water becomes cloudy with blood), making vision difficult. Other organisms, such as the knifefish, take the electrical sense further, generating their own stronger electrical currents and using these to "probe" their surroundings. The electrical current is generated by modified muscle cells, and for the system to work properly, depends on the body being kept in a straight line, thus forming the selective pressure for the unusual form of swimming exhibited by the knifefish, which uses its anal fin for propulsion, eliminating the need to bend the spine. Still other electric fish go further still; after locating prey with the electric sense they generate a powerful current to stun or kill the prey; you are no doubt familiar with electric catfish, eels, and rays.
At very close range, the sense of touch may be very important. This is particularly true of invertebrates, although it is of course used by vertebrates as well. Some damselfly larvae, for instance, locate their prey visually and use strong stereoscopic vision to maneuver into the proper position. The final lunge in some species, however, is controlled by the antennae, which are used as range finders. When the prey is in contact with both antennae, the range and position are ideal, and the antennae spring back out of the way, allowing the labium to spring forward and capture the prey (Fig. 3). Stonefly larvae also play their antennae over their prey before lunging. Apparently they get both directional and other information from this. An anti-predatory device employed by some mayflies seems to work to fool stoneflies; when touched by the antennae of a stonefly; the ephemerellid mayfly's response is to bend its tail forward over its back in a "scorpion posture". This apparently fools the stonefly into thinking that the mayfly is too big to eat. When it come to finding prey by touch, however, the octopus is king, using its long tentacles to explore crevices and capture its prey.
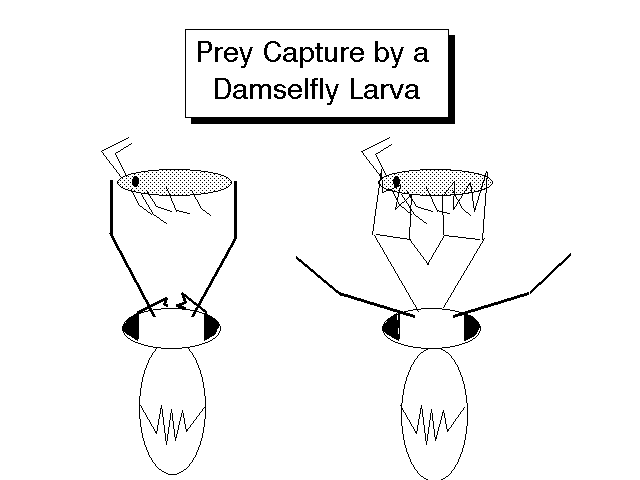
Figure 3. Capture of an amphipod by a damselfly larva. When the amphipod is detected between the two antennae (above, left), the antennae spring back and the labium is shot forward to grasp the prey (above, right). The labium is held folded under the head when not in use. In the images below, you can see the extent to which the labium can be extended in the damselfly Archilestes grandis.

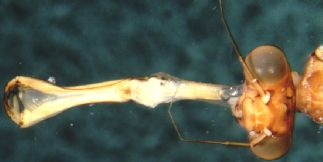
Capturing prey often involves catching highly mobile prey, and a number of aquatic predators are very swift. We term predators that actively search out mobile prey as pursuit predators. These organisms seek out their prey, which may in turn also be quick. Pursuit predators are thus distinguished by being fast and having good distance senses; common examples would be sharks. There are other strategies, however. Searchers are highly mobile themselves, but search out sedentary prey. Examples here might include many starfish; and it is really a minor distinction between searcher-predators and searcher-herbivores, which also must seek out a sedentary food source. Search strategies employed by both predators and herbivores are currently a "hot" area of research. Ambushers take a different tack. They are sedentary, and lie in wait for mobile prey. Many dangle a lure of some sort to draw in the unwary. Alligator snapping turtles, for instance, dangle a worm-like extrusion from their tongues. Perhaps the best in this vein, however, are certain freshwater mussels that modify a portion of their mantle to resemble the females of certain species of fish (Figure 4). When an unsuspecting male fish approaches, the mussel squirts out a cloud of larvae, which attach themselves as ectoparasites on the gills of the fish (Figure 5).

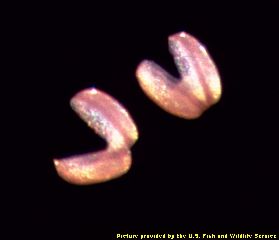
Figure 4 (left) Mantle of a mussel adapted to resemble a fish. Figure 5 (right) Glochidia (larvae) of a mussel. These organisms clamp down on the gills or fins of a fish, and draw nutrients as ectoparasites on the fish. They drop off after a few weeks, and usually do not seriously injure the fish.
Prey may be captured by use of a number of structures. Many organisms simply engulf their prey with the mouth. Fish and turtles are especially good at this; the prey is overcome through a combination of lunge and suction. The seahorse and its relative the pipefish also use suction to capture their prey. Other organisms use tentacles or other appendages to seize prey. Among the latter the preying-mantis like forelegs of the mantid shrimps are particularly interesting. Adapted variously to slice, crack, or snatch prey, they are among the fastest moving structures in water. One species, which uses knobs on the ends of the forelegs to crack open mollusk shells, can hit with an impact equivalent to a .22 caliber bullet (they can and have cracked aquarium glass, making them fun to tease at major aquariums). Dragonflies and damselflies use an extensible labium to capture prey.
Once captured, the prey must be overcome. In many instances, this is simply a matter of bolting it down, that is, swallowing it whole and alive, and allowing the digestive juices to go to work. In other cases, various structures are used to tear or cut up the prey before it is swallowed. Octopuses and squids have beaks that tear prey and inject a poison. All marine snakes are poisonous as well, injecting a poison similar to that of the cobras. Arthropods generally have tearing or cutting structures associated with the legs; examples would include the chelipeds of lobsters and crayfish, or the gnathobases of horseshoe crabs. Starfish that prey on bivalve mollusks wrap themselves around the clams and pry it open just enough to evert their stomach into the shell cavity, where it excretes a poison and digestive juices that digest the unlucky mollusk within its own shell. Mollusks are not defenseless, however; cone-shell gastropods are quick enough to lunge at small fish and overcome them with a toxin, and some snails use their radula to bore through the hard outer cases of barnacles and bivalves.
Suctorial feeding, where the juices of the prey are sucked out, is also common in water. Aquatic insects in the order Hemiptera, such as water striders, water boatmen, giant water bugs, and water scorpions, all feed in this manner. Likewise, leeches and lampreys latch onto their prey and suck out the juices. This type of predation is often called ectoparasitism. The whole phenomenon of parasitism can be considered as a subset of predation. To distinguish a predator from a parasite, several things can be considered, but remember, as in all continua, the lines separating these two modes of life often blur. Usually, but not always, predators tend to be bigger than their prey, while parasites tend to be smaller than their hosts are. Predators almost always kill from the outside and engulf their prey; parasites may be external or internal, and usually do not directly ingest host tissue other than blood. Many parasites simply live in the guts of their hosts and absorb food from the surrounding fluids. Parasites often have degenerate systems, especially sensory systems, mouthparts, and digestive systems, and parasites often show complicated life cycles with alternating hosts and an emphasis on reproduction. Predators almost always kill their prey in the act of predation, while a "good" parasite usually does not kill its host directly, although it may weaken it or leave it open to infection. Parasitoids are insect endoparasites, usually of other insects, that differ from other parasites in that they usually do kill their prey. Insects can get away with this because they reproduce so rapidly. In aquatic systems there are some tiny wasps which "fly" underwater to parasitize other insects. Both hunters and parasites consume high-quality food and thus have simple, short guts.
Grazers and Browsers:
As already mentioned, these organisms really act much like a searching-type predator in that they are mobile and search for a sessile "prey" (in fact, many organisms in this group really don't distinguish between plant and animal food - whatever they can shovel in their mouths). The common problems they face are finding the patches of food and removing it from the substrate.
Many herbivores are highly mobile swimmers with great maneuverability. They must be able to "hover" over a site while removing food, and often must get into tight spaces where algae may be attached. Other herbivores crawl over the bottom to patches of algae. Once the food is found, hard structures must be brought into play as has already been discussed. Even at this point, however, these organisms still have formidable obstacles to overcome. Many plants defend themselves with poisonous chemicals and indigestible bulk such as cellulose that makes them nutritionally unappealing. Herbivores may counter by careful, selective feeding on only those plants with the fewest chemical defenses, with elongate guts so that material may be digested better, and with symbionts bearing the necessary enzymes to digest cellulose and other materials that the herbivore cannot digest on its own. There is a constant evolutionary "arms race" as herbivores try to overcome the new defenses that are always arising among plants.
Avoiding Predation
The evolutionary war between plants and herbivores is also being fought between prey (both herbivores and carnivores) and their predators. Any potential prey will try to avoid being eaten, and the predators must overcome these defenses. Basically, there are three main tactics the prey can try - to increase search time, increase handling time, or actively repelling the predator. The first two rely on some economic-ecological principles, the first being that time is like money (economics) or energy (ecology). If the predator requires too much time to successfully capture and eat a prey item, it will leave that species alone in the future. It doesn't do the individual any good, but it may help out that individual's offspring.
Increasing search time is another way of saying hiding. You are no doubt aware of the cryptic patterns exhibited by organisms that makes them hard to see. These organisms often have various protuberances to help break up their body outline, and are colored to resemble their surroundings. Some of the best examples are the various organisms found in the drifting seaweed Sargassum. Some organisms take a slightly more active role; the decorator crab actively finds anemones and other sessile organisms and encourages them to grow on its shell, helping to camouflage it (those stinging tentacles don't hurt either!). Stripes and fake eyespots on many aquatic organisms also help to confuse predators and divert their attacks away from critical areas.
Increasing handling time can be done in a number of ways. Schooling fish are easily found but hard to capture since their numbers confuse predators. Shells, especially those with ridges or spines that effectively increase their size without increasing the weight proportionately, make it tough for a predator to get to the nutritious parts of the animal. There is often a close correspondence between the size of the shell and the size of the predominate predators' jaws (claws, gnathobases, whatever).
Active defenses include attacking the predator directly, either alone or in concert with others of your species. Predators, of course, have the necessary weapons at hand for such attacks, but many herbivores can improvise effectively. Running is a type of active defense, and it is often enhanced by trickery, such as leaving a severed (but replaceable) limb to distract the predator, or squirting a cloud of ink or other chemicals in its path to confuse or irritate it. Other means of active chemical defense may be widespread in aquatic systems, but if they are, they are poorly known as compared to the myriad of chemical defenses exhibited on land. Many examples of chemical defenses are known from insects, perhaps it is because the insects are not one of the dominant aquatic taxa that we do not see more examples of chemical defenses in aquatic systems.
Microhabitats and Feeding
Often, the microhabitat that an organism inhabits is closely tied to its feeding. Scrapers will be found on top of rocks where algae grow, while deposit feeders will be found in eddies near the bottom where detritus settles out. Filter feeders will gather where the currents are strong, and predators will be found everywhere. The two figures below (Figs. 6 & 7) illustrate somewhat the complex environment that develops in aquatic systems.
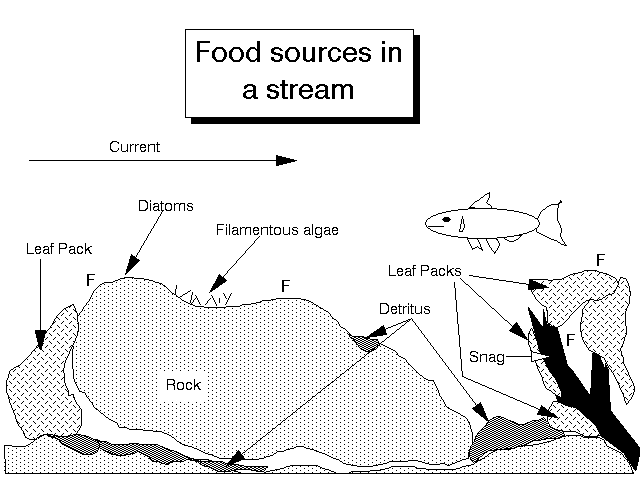
Figure 6. Microhabitats in a stream. Scrapers will be found on the open rock faces, while filters move to areas where they can intercept the current (F). Deposit feeders will be found in the crevices under the rocks and among the detritus. Shredders will occupy the leaf packs, and numerous detritivores will inhabit burrows in the bottom sediments. Miners may inhabit the wood of the snag. Predators will be found (in lesser numbers) in all these microhabitats. Most of the niches are filled by aquatic insects with some representation by other phyla.

Figure 7.. Food sources on a coral reef. Photosynthesis occurs on all exposed surfaces, and many of the organisms such as sponges, sea fans, coral, and anemones also house photosynthetic endosymbionts. As in the stream, detritivores of various types live under rocks and coral and come out to feed on deposits. A number of organisms are adapted to either filter out material from the water (sponges, corals, sea fans), or intercept it before it is deposited (crinoids). Predators are found throughout the system. As opposed to freshwater systems, where fish are primarily predators, fish on a coral reef fill a variety of niches, and other niches are filled by members of various phyla, particularly annelids, crustaceans, echinoderms, and mollusks.
Further Reading
McShaffrey, D. 1992. Comparative functional morphology of larval
Stenacron interpunctatum and Rhithrogena pellucida (Ephemeroptera: Heptageniidae) and
Ephemerella needhami (Ephemeroptera: Ephemerellidae) with applications in mayfly taxonomy and ecology. Proceedings of the VII International Conference on
Ephemeroptera.
- Barnes, R.S.K., P. Calow, and P.J.W. Olive. 1988. The invertebrates: a new synthesis. Blackwell Scientific Publications. Oxford. 582pp. Read Chapter 9
- Craig, D.A. 1990. Behavioral hydrodynamics of Cloeon dipterum larvae (Ephemeroptera: Baetidae). Journal of the North American Benthological Society. 9:346-357. Read article, particularly noting relationship between antennae and the boundary layer.
- Ehrlich, P.R. and Roughgarden, J. 1987. The Science of Ecology. MacMillan Pub. Co., New York. 710 pp. Review information on guilds and trophic levels
- Lissmann, H.W. March, 1963. Electric Location by Fishes. Scientific American. Read article.
- McCafferty, W.P. 1981. Aquatic Entomology Science Books Intl., Boston. 448 pp. Read Chapter 3, pp. 35-39.
- McShaffrey, D. and W.P. McCafferty. 1987. The behavior and form of Psephenus herricki (DeKay) (Coleoptera: Psephenidae) in relation to water flow. Freshwater Biology. 18:319-324.
- McShaffrey, D. & McCafferty, W.P. 1988. Feeding behavior of Rhithrogena pellucida (Ephemeroptera: Heptageniidae). J. North Amer. Benthol. Soc. 7: 87-99.
- Merritt, R.W. and K.W. Cummins (eds.). 1978. An introduction to the aquatic insect of North America. Kendall/Hunt, Dubuque. 441 pp.
- Peckarsky, B.L. and R.S. Wilcox. 1989. Stonefly nymphs use hydrodynamic cues to discriminate between prey. Oecologia 79:265-270. Read Article
- Pough, F.H, J.B. Heiser, and W.N. McFarland. 1989. Vertebrate Life, 3rd Edition. MacMillan Publishing Co., New York. 943 pp.Read pp. 130-139, 282-289.
- Rubenstein, D.I. and M.A.R. Koehl. 1977. The mechanisms of filter feeding: some theoretical considerations. American Naturalist 111:981-994.
Supplemental Bibliography - Feeding
- Brown, D.S. 1960. The ingestion and digestion of algae by Cloeon dipterum L. (Ephemeroptera). Hydrobiologia 16: 81-96.
- Chance, M.M. 1970. The functional morphology of the mouthparts of black fly larvae (Diptera: Simuliidae). Quaestiones Entomologicae 6: 245-284.
- Clifford, H.F. 1982. Life cycles of Mayflies (Ephemeroptera), with special reference to voltinism. Quaestiones Entomologicae 18:15-90.
- Cummins, K.W. 1973. Trophic relations of aquatic insects. Annual Review of Entomology 18: 183-206.
- Cummins. K.W. and M.J. Klug. Feeding ecology of stream invertebrates. Annual Review of Ecology and Systematics 10: 147-172.
- Cummins, K.W., G.F. Edmunds, and R.W. Merritt. 1984. Summary of ecological and distributional data for Ephemeroptera (mayflies). Table 10A, Pages 122-125. in R.W Merritt and K.W. Cummins (editors). An introduction to the aquatic insects of North America. Kendall-Hunt Publishing Company, Dubuque, Iowa.
- Cummins, K.W., and R.W. Merritt. 1984. Ecology and distribution of aquatic insects. Pages 59-65 in R.W. Merritt and K.W. Cummins (editors). An introduction to the aquatic insects of North America. Kendall-Hunt Publishing Company, Dubuque, Iowa.
- Dahl, C., L.E. Widahl, and C. Nilsson. 1988. Functional analysis of the suspension feeding system in mosquitoes (Diptera: Culicidae). Annals of the Entomological Society of America 81:105-127.
- Devitt, B.D., and J.J.B. Smith 1985. Action of mouthparts during feeding in the dark-sided cutworm,Euxoa messoria (Lepidoptera:Noctuidae). Canadian Entomologist 117: 343-349.
- Flowers, R.W., and W.L. Hilsenhoff. 1978. Life cycles and habitats of Wisconsin Heptageniidae (Ephemeroptera). Hydrobiologia 60: 159-171.
- Froehlich, C.G. 1964. The feeding apparatus of the nymph of Arthroplea cogener Bengtsson (Ephemeroptera). Opuscula Entomologica 29: 188-208.
- Keltner, J., and W.P. McCafferty 1986. Functional morphology of burrowing in the mayflies Hexagenia limbata and Pentagenia vittigera. Zoological Journal of the Linnaean Society 87:139-162.
- Lamp, W.O., and N.W. Britt 1981. Resource partitioning by two species of stream mayflies (Ephemeroptera: Heptageniidae). Great Lakes Entomologist 14: 151-157.
- McCafferty, W.P. and B.L. Huff, Jr. 1978. The life cycle of the mayfly Stenacron interpunctatum (Ephemeroptera: Heptageniidae). Great Lakes Entomologist 11:209-216.
- McShaffrey, D. and W.P. McCafferty. 1986. Feeding behavior of Stenacron interpunctatum (Ephemeroptera: Heptageniidae). Journal of the North American Benthological Society 5:200-210.
- Pucat, A.M. 1965. The functional morphology of the mouthparts of some mosquito larvae. Quaestiones Entomologicae 1: 41-86.
- Strenger, A. 1953. Zur Kopfmorphologie der Ephemeridenlarven. I. Teil Ecdyonurus und Rhithrogena. Osterreichische Zoologische Zeitschrift 4: 191-228.
- Trueman, E.R. 1968. A comparative account of the burrowing process of species of Mactra and of other bivalves. Proceedings of the Malacological Society of London 38:139-151.
- Wiley, M.J., and S.L. Kohler 1980. Positioning changes of mayfly nymphs due to behavioral regulation of oxygen consumption. Canadian Journal of Zoology 58: 618-622.
- Wodsedalek, J.E. 1912. Natural history and general behavior of the Ephemeridae nymphs Heptagenia interpunctata. Annals of the Entomological Society of America 5: 31-40.